Inmunidad digestiva del lechón lactante y destetado
17-nov-2005 (hace 19 años 4 meses 14 días)
Las enfermedades entéricas, y en particular las diarreas de causa infecciosa, representan uno de los problemas más comunes en los cerdos desde su nacimiento hasta la fase de engorde. La aparición de estas enfermedades se genera por la interacción de tres elementos clave: la presencia de patógenos entéricos en la explotación, las condiciones de higiene y manejo y la inmunidad de los cerdos frente a los patógenos circulantes en la granja. Este último aspecto resulta crítico y de especial importancia en las diarreas de los lechones lactantes y en el destete.
La inmunidad en el tracto digestivo es especialmente compleja y conjuga defensas de tipo natural o inespecífico con mecanismos específicos (anticuerpos e inmunidad celular). Dentro de los mecanismos naturales cabe destacar el pH ácido del estómago que destruye una gran cantidad de patógenos. Sin embargo, la acidez del contenido gástrico puede verse modificada por la dieta o por la edad, como ocurre en los lechones lactantes de pocos días cuyo pH es menos ácido y; por lo tanto, tiene un menor efecto de protección. Una segunda barrera física la constituye la flora saprofita presente en el intestino. Por una parte, la flora saprofita ocupa un nicho ecológico que impide el asentamiento de otra flora patógena y, por otra parte, los metabolitos de algunas de las bacterias saprofitas pueden actuar como inhibidores del crecimiento de determinados patógenos. Un ejemplo de este último hecho es la producción de ácidos grasos volátiles por lactobacilos presentes en el intestino.
A nivel práctico, el intestino representa un órgano linfoide que se organiza en una serie de folículos linfoides a los que se denomina placas de Peyer (fundamentalmente compuestas por células B), más una serie de linfocitos presentes en la lamina propia y el epitelio intestinal (linfocitos intraepiteliales). Otro componente importante de la inmunidad intestinal son las células M, cuya función principal es la de capturar, mediante endocitosis, proteínas y péptidos antigénicos que el sistema inmunitario se encargará de reconocer como provenientes de patógenos o de los alimentos. En el primer caso se desarrollará una respuesta activa de defensa mientras que en el segundo caso se desarrollará tolerancia o se obtendrá una respuesta anérgica (figura 1).
Figura 1. El intestino actúa de hecho como un órgano linfoide. Por una parte existen folículos linfoides denominados placas de Peyer, formados fundamentalmente por células B. Por otra parte, a lo largo del intestino se presenta células linfoides intraepiteliales y mucosales.
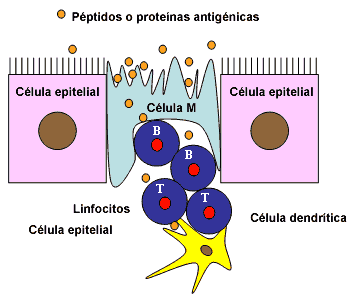
En el epitelio intestinal existe un tipo particular de células denominado células M, cuya función es la captación de antígenos presentes en el intestino. Estas células M están en contacto estrecho con los linfocitos B y T y con células dendríticas que actúan de presentadoras de antígeno para los linfocitos.
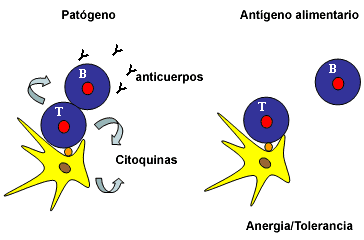
Si el antígeno captado corresponde a un patógeno (izquierda), se activan los mecanismos de respuesta inmunitaria específica. Entre estos mecanismos destaca la producción de anticuerpos (mayoritariamente IgAs) y la secreción de citoquinas que regularán dicha respuesta. Si se trata de un antígeno alimentario, la respuesta inmune no se produce (anergia) o se desarrolla tolerancia; es decir, el antígeno alimentario no se reconoce como extraño al organismo.
La respuesta inmune activa frente a patógenos entéricos en su gran mayoría depende de la producción de anticuerpos a nivel mucosal. Como acabamos de ver, el intestino actúa como un órgano linfoide en el cual puede producirse el reconocimiento de los antígenos presentes. Esta es la razón por la que la mayoría de vacunas efectivas contra patógenos entéricos que se han desarrollado, tanto en el cerdo como en otras especies, se administran por vía oral. En muchos casos, la administración parenteral de la misma vacuna consigue el desarrollo de una inmunidad sistémica pero es poco efectiva para inducir inmunidad en las mucosas digestivas.
Otro punto importante a considerar en la inmunidad intestinal de los lechones es la transferencia de anticuerpos a través del calostro y la leche. El intestino sólo es permeable al paso de inmunoglobulinas hacia la sangre durante las primeras 24-36 horas desde el nacimiento del lechón. Sin embargo, el intestino posee receptores para las inmunoglobulinas de tipo A (IgA) lo que permite que estos anticuerpos queden retenidos en el intestino mientras el lechón recibe aporte de leche materna. La presencia de estas IgA en el intestino permite que el lechón tenga protección frente a los patógenos entéricos a los que su madre hubiera desarrollado anticuerpos (figura 2). Por ello resulta crucial una correcta adaptación e inmunización de las cerdas frente a los patógenos entéricos comunes de la explotación. En este mismo concepto está basada la inmunización por retroalimentación. Por ejemplo, ante un brote de gastroenteritis transmisible, una de las formas de controlar la infección en las parideras es contaminar el alimento de las cerdas gestantes con heces de animales infectados. La cerda se infectará sufriendo la enfermedad de forma suave o subclínica pero desarrollará inmunidad mucosal. Los linfocitos B específicos del virus migrarán a la glándula mamaria y las inmunoglobulinas se secretarán en la leche y protegerán al lechón. Aunque el método es efectivo debe considerarse como un último recurso ya que la infección de las cerdas con heces de otros animales puede servir también como método de diseminar otros patógenos en la explotación y, en consecuencia, tener efectos indeseables. Una vez el lechón ha sido destetado, estas IgA retenidas en el intestino desaparecen paulatinamente y, alrededor de una semana post-destete, su efecto protector ha desaparecido. Este es uno de los motivos por los que es muy frecuente que aparezcan problemas diarreicos en esa época.
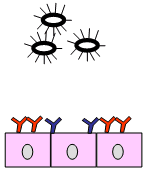
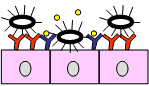
Figura 2. Protección lactogénica conferida por las IgA frente a cepas enterotoxigénicas de E. coli.
| E. coli enterotoxigénico | |
 |
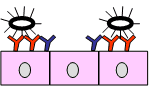 |
| Epitelio intestinal con IgAs lactogénicas específicas de las fimbrias (rojo) o de las toxinas (azul) | Las IgA neutralizan las fimbrias e impiden la adhesión de la cepa de E. coli |
 |
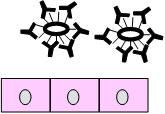 |
| Si alguna bacteria llega a adherirse y produce enterotoxina, las IgA específicas la neutralizan | Las bacterias neutralizadas son eliminadas y las IgA se pierden |