El papel del tejido magro en el manejo de las reservas corporales
07-feb-2011 (hace 14 años 2 meses 19 días)
Es bien conocido que el mantenimiento de la condición corporal en cerdas reproductoras es esencial para garantizar unos buenos rendimientos productivos a lo largo de su vida. También es bien conocido que las cerdas de hoy en día cuentan con un menor nivel de reservas grasas que las cerdas de hace 20-30 años para hacer frente a unas exigencias reproductivas muy elevadas (mayor número de lechones nacidos y destetados, lactaciones cada vez más largas,…). La capacidad de las reservas grasas para hacer frente a situaciones de más demanda energética es escasa así que cuando no conseguimos equilibrar la alimentación de estas cerdas con sus necesidades, que es lo que generalmente ocurre en lactación, éstas tienden a perder peso y reservas.
Estas pérdidas de peso se componen, generalmente, de una cantidad máxima de tejido graso que se pierde en primer lugar seguido de una cantidad de tejido magro que sigue a la pérdida grasa. Al tener menores cantidades de reservas grasas, la pérdida de masa proteica es muy frecuente en las cerdas de hoy en día. Cuando la alimentación se maneja adecuadamente, las pérdidas de peso en lactación no deberían ser mayores de unos 10 kg (Noblet et al., 1990; Close and Cole, 2001) y deberían poder ser totalmente recuperadas a lo largo de la siguiente gestación.
En términos de recuperación de reservas, cierto es que las reservas proteicas son más difícilmente recuperables a través de la alimentación que las reservas grasas ya que las reservas magras son parte “estructural” de los animales, sobre todo en cerdas primerizas. Esta puede ser la causa de que animales con una condición corporal muy perjudicada al final de la lactación, y, por tanto, con una elevada pérdida de tejido magro sean muy difícilmente recuperables en ciclos siguientes. En este sentido varios estudios indican que los genotipos magros actuales dependen más de la masa proteica que los genotipos anteriores para mantener la función reproductora (Cia et al., 1998; Sinclair et al., 1998). Hace ya algunos años, King (1987) sugirió que una excesiva movilización de proteína durante la lactación era más perjudicial para la fertilidad que una excesiva movilización de grasa. Posteriormente, algunos autores han observado la importancia de la conservación de la masa muscular sobre el tamaño de la siguiente camada en cerdas primíparas. Touchette et al., (1998) observaron que pérdidas de masa muscular durante la lactación superiores al 10% resultaban en una disminución de la productividad numérica de primerizas en el segundo ciclo. También Clowes et al. (2003a) han asociado una reducción en la función ovárica muy marcada y una reducción de los rendimientos en lactación (producción de leche y crecimiento de la camada) con pérdidas de proteína corporal de un 12% o superiores.
Además de las pérdidas de tejido magro en lactación también se ha visto que el nivel de reservas magras en el momento del parto es importante para minimizar los problemas post-destete. En este sentido, cerdas con mayores niveles de reservas magras al parto conseguidas mediante una alimentación especializada en gestación (piensos con mayores niveles de proteína y lisina) presentaban una mejor función ovárica y rendimiento en lactación que las cerdas con menores niveles de reservas (Clowes et al., 2003b). Sin embargo, aunque la cantidad de masa proteica perdida parece ser muy importante para la posterior recuperación de la cerda, el mantenimiento de un cierto nivel de grasa sigue siendo crucial en la función reproductiva. En este sentido, Whittemore (1996) sugirió que si el cociente lípidos:proteína en cerdas primíparas era inferior a 1:1, la función reproductiva se veía perjudicada.
En definitiva, una excesiva pérdida de peso durante la lactación tiene consecuencias negativas sobre la reproducción mediadas, muy probablemente, por la pérdida de reservas grasas y magras y por el estado metabólico que esto genera en el animal. En este sentido, estudios recientes (Cebrian et al., 2011) indican que es posible que las cerdas jóvenes de hoy en día sean más sensibles a perder tejido magro si la alimentación no es adecuada (nivel de alimentación, composición del pienso) (figura 1) y que debamos plantearnos la formulación de piensos con diferentes niveles de aminoácidos en las cerdas jóvenes actuales (mayores niveles), haciendo hincapié en los periodos de recuperación de reservas como la gestación.
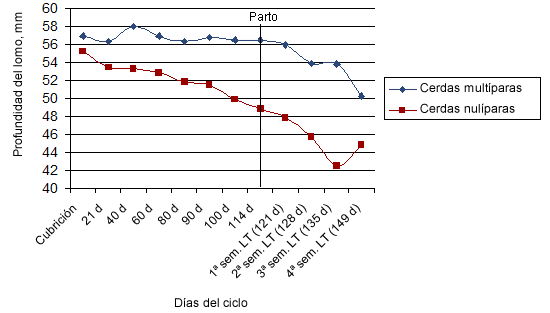
Figura 1. Evolución de los niveles de profundidad de lomo durante un ciclo productivo en cerdas primerizas y multíparas.
En la práctica, no existen métodos directos para medir las reservas magras a nivel de campo. A nivel experimental se emplean generalmente equipos de ecografía mediante los cuales se mide la profundidad de lomo (PL) como índice de reservas magras (al igual que se mide el espesor de grasa dorsal (EGD) como indicador del nivel de reservas grasas) (figura 2). Sin embargo, en las nuevas genéticas magras se ha comprobado que la nota de condición corporal (CC) comúnmente utilizada en granja para valorar el nivel de EGD (r = 0,55) se encuentra también relacionada con la PL (r = 0,49) y el peso vivo (r = 0,45) del animal, es decir, que además del estado de reservas grasas mide también la conformación del animal (Cerisuelo et al., 2005). Por lo que, indirectamente, el método visual es el único método práctico capaz hoy en día de valorar las reservas magras en un núcleo de reproductoras. En este sentido quizá sería conveniente adaptar los patrones de medición de la nota de CC, enfocados hasta el momento hacia la búsqueda de depósitos grasos, hacia una valoración más estricta de la conformación del animal.
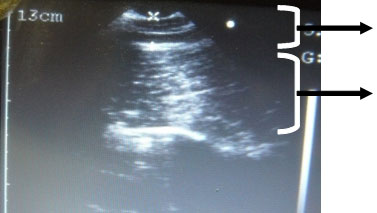 |
Espesor de grasa dorsal |
| Profundidad de lomo | |
Figura 2. Imagen ecográfica del espesor de tocino dorsal y la profundidad de lomo de una cerda.