

El catabolismo de la treonina en los cerdos puede producirse a través de dos rutas metabólicas (figura 1): L-treonina deshidrogenasa y treonina deshidratasa. En el primer caso el producto final es glicina y en el segundo ácido ketobutírico, el cual puede convertirse en propionil CoA. La vía treonina deshidrogenasa es la más importante y mayoritaria. La degradación de la treonina es constante y bastante baja cuando los aportes son inferiores a las necesidades, aumentando considerablemente cuando se suplementa por encima de las necesidades. Sin embargo, cuando el exceso es importante, la actividad de las rutas metabólicas de degradación es insuficiente produciéndose un acumulo de treonina en el pool corporal afectando negativamente al apetito.
La gran importancia de la treonina en las funciones de mantenimiento se explica por su alta utilización en el tracto gastrointestinal y sus órganos asociados (bazo, páncreas e hígado), representado aproximadamente un 55 % de la treonina de la dieta. Esta utilización intensiva de la treonina de la dieta en el tracto gastrointestinal incluye en una pequeña proporción el catabolismo, principalmente en el hígado y en menor medida en el páncreas, y en una gran proporción la síntesis de proteínas del intestino. Se ha descrito la gran importancia de la treonina en la síntesis de mucina (relación treonina/lisina del 713%; figura 2) y en otras secreciones intestinales endógenas (treonina/lisina de 142% en las pérdidas endógenas). La utilización de treonina para la síntesis de proteínas digestivas se verá afectada por la ingestión de materia seca, el tamaño del tracto digestivo o por características de la dieta como por ejemplo la presencia de componentes indigestibles. Así, varios autores han encontrado una reducción en la secreción de mucina y en el peso del intestino al emplear dietas claramente deficientes en treonina, aumentando además la incidencia de diarreas no infecciosas.
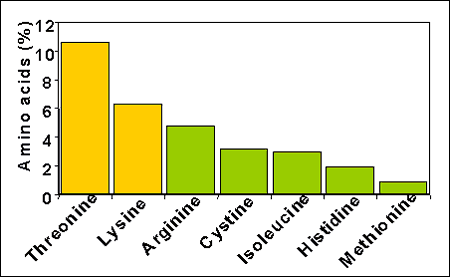
Investigaciones recientes muestran la importancia de la treonina en la respuesta inmunitaria, siendo las inmunoglobulinas y las proteínas de fase aguda ricas en este aminoácido (157% respecto a la lisina en el caso de las IgG; figura 3). Se han descrito respuestas de la suplementación de la dieta con treonina sobre la producción de IgG tanto en cerdas a nivel sérico y en leche, como en cerdos de cebo en respuesta a la vacunación.
Sin embargo, todas las utilizaciones metabólicas de mantenimiento descritas explican sólo en parte la baja eficacia de deposición de la treonina en la proteína corporal, existiendo la posibilidad de la presencia de otras rutas metabólicas de catabolismo aun no identificadas y siendo necesario estudiar el impacto de la flora intestinal sobre la disponibilidad de la treonina para el animal.
A nivel práctico se deben considerar las necesidades de treonina para la deposición de proteína crecimiento corporal. Sin embargo, debido a su gran importancia en las funciones de mantenimiento principalmente relacionadas con el sistema digestivo e inmunitario, sería de interés considerar el incremento de su suplementación o de su relación respecto a la lisina en situaciones de adaptación digestiva y de alta exposición del sistema inmune.
Figura 1. Vías metabólicas de catabolismo de la treonina (de Blas et al., 2000)
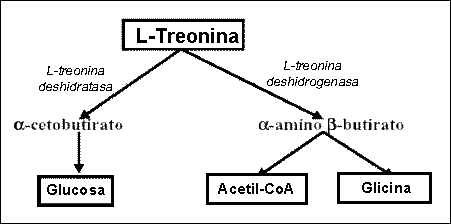 |
Figura 2. Composición aminoacídica de las mucinas (Ball, 2002)
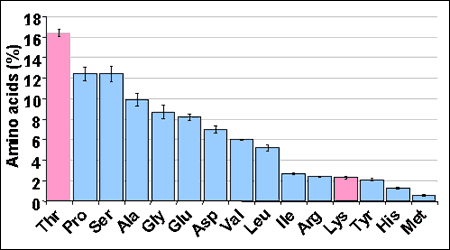 |
Figura 3. Composición aminoacídica de las inmunoglobinas G (Bowland 1966; Han and lee, 2000)
 |
Así lo ve Imasde
