El sistema inmunitario y la inmunidad en el cerdo: la piel
09-abr-2018 (hace 7 años 10 días)
Representa un sistema coordinado en el que la barrera física, compuesta por el epitelio, las células inmunes residentes, el microambiente local y su microbioma, trabajan juntos para mantener la homeostasis del cuerpo.
La piel está compuesta por tres capas principales y por anexos que incluyen folículos pilosos y glándulas sebáceas y sudoríparas (figura 1):
- La epidermis (20-26 μm de grosor), primera barrera externa, donde la descamación continua de queratinocitos, la desecación, el pH bajo, el sebo, los ácidos grasos y las bacterias comensales (hasta 1012microorganismos/m2), garantizan su integridad y dificultan la penetración por parte de los patógenos;
- La dermis compuesta por unos pocos fibroblastos, fibras de colágeno y elastina, contiene sangre, vasos linfáticos y células del sistema inmune innato y adquirido;
- El subcutis, compuesto por una capa grasa subepidérmica, con algunos vasos sanguíneos y linfáticos;
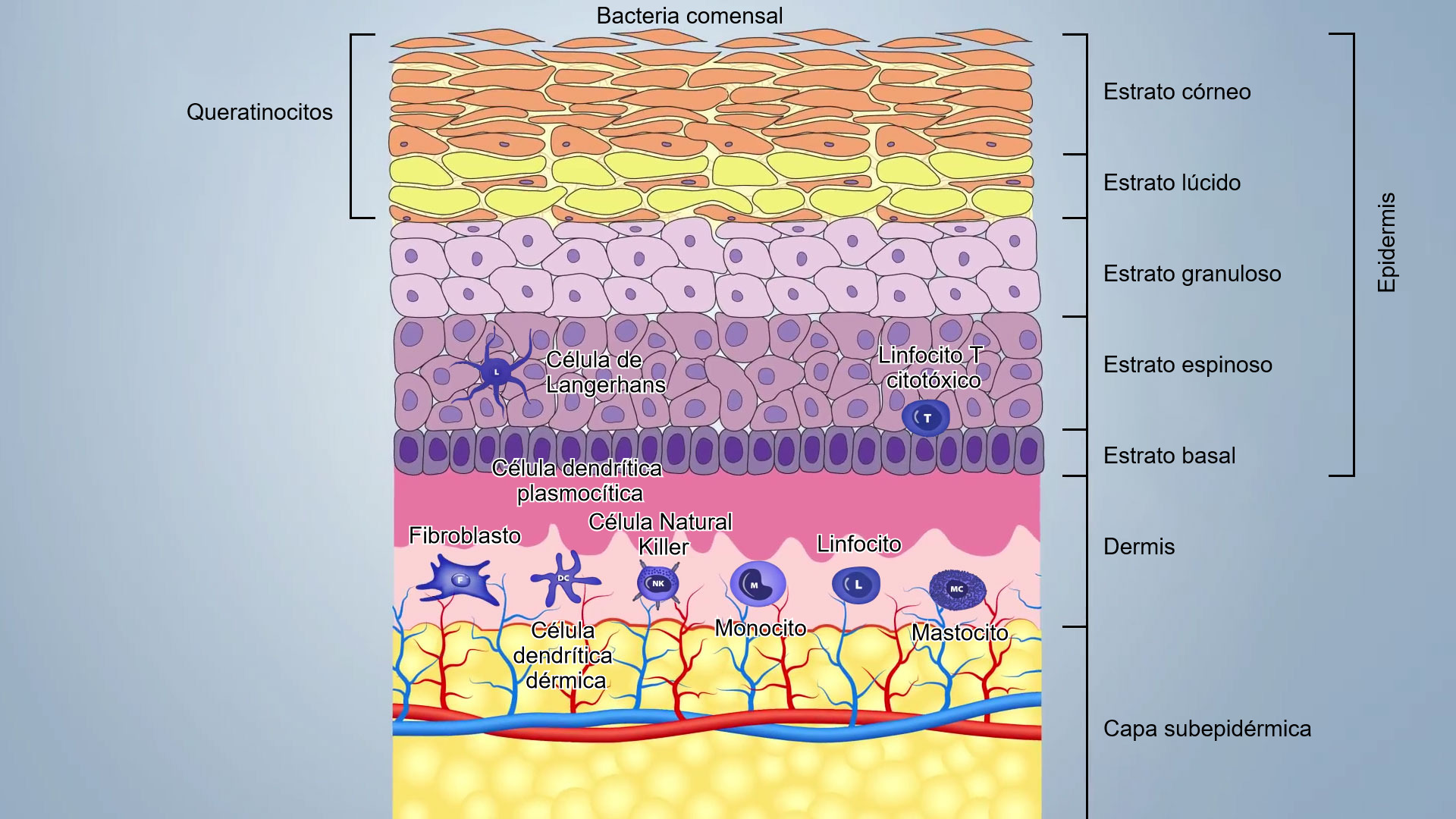
Estas capas están estrechamente unidas y forman una barrera que varía de 50 a 120 μm, dependiendo de la ubicación en el cuerpo: en la ingle y las axilas son más delgadas que en las otras partes del cuerpo.
Componentes inmunológicos de la epidermis
Los queratinocitos son las primeras células defensivas que expresan una amplia gama de PRR (Pathogen Recognition Receptors) como TLR (TLR-1, TLR-2, TLR-4, TLR-5 y TLR-6) y lectinas de tipo c; también secretan una amplia gama de citoquinas proinflamatorias y péptidos antimicrobianos (AMP). De hecho, los queratinocitos pueden reconocer PAMP (Pathogen Molecular Pattern) mediante PPR y desencadenar inflamación secretando interleucinas (IL-1), IFN, citoquinas, factores de crecimiento, quimiocinas, defensinas. También pueden expresar MHCII y presentar antígenos.
El antígeno atrapado en la superficie es capturado e internalizado por las células de Langerhans (CL), un subconjunto especial de células dendríticas epidérmicas localizadas entre los queratinocitos de la capa basal y el estrato espinoso. Las CL procesan el antígeno y lo presentan a las células T auxiliares cercanas. En la epidermis porcina, las CL tienen una morfología típica de células dendríticas (CD) y expresan moléculas de MHCII y moléculas de superficie como CD1, CD172 y CD207.
Las CL epidérmicas también son necesarias para mantener una respuesta citotóxica efectiva y para la diferenciación de Th17 desencadenada por la secreción de IL-6, IL-1β e IL-23. En el cerdo, el 50-70% de las células dendríticas de la piel son CL.
La epidermis porcina también contiene un pequeño número de células γδT, la mayoría de las cuales son células T de memoria citotóxica a largo plazo, residentes en la piel. Algunas de estas células, cuando se activan, pueden matar directamente a los queratinocitos infectados y reclutar otras células.
Las inmunoglobulinas pueden atravesar la piel mediante transudación y las IgG1, IgG2 e IgA también pueden sintetizarse localmente.
Vídeo 1. Respuesta inmune de la piel.
Componentes inmunológicos de la dermis
Las células dendríticas CD1+ representan las células inmunes dominantes y más importantes de la dermis. Un subconjunto de estas células (células dendríticas dérmicas) puede presentar el antígeno e inducir la respuesta primaria de las células T en el ganglio linfático, mientras que otro subconjunto (células dendríticas plasmocitoides) se especializa en la producción de una gran cantidad de interferón tipo I en respuesta al reconocimiento de ácidos nucleicos virales.
La dermis contiene una gran cantidad de linfocitos que se ha estimado que es incluso superior al número de células en la circulación sanguínea. Las células T están dirigidas a la dermis por el antígeno leucocitario cutáneo (CLA- cutaneous leukocyte antigen). Los principales subconjuntos de células T son CD4+, Th1, Th2 y Th17.
Se ha identificado un subconjunto de células T circulantes, que se dirigen a la piel y producen IL-22, siendo capaces de mantener la función de barrera en superficies expuestas del cuerpo.
Los macrófagos (CD14+) son raros en la dermis, pero los monocitos,que se diferencian rápidamente en macrófagos, se reclutan eficazmente después de que se produzca una lesión y la inflamación.
En la dermis también hay células natural killers (NK); eliminan células infectadas por virus y células cancerosas con una citotoxicidad no restringida por MHC (Major histocompatibility complex). Las NK se activan por interferones u otras citoquinas liberadas por macrófagos o queratinocitos. Las células NK expresan receptores inhibidores (KIR- killer cell immunoglobuline-like receptor) que reconocen el MHC-I y evitan ataques indeseados sobre el propio organismo. Pueden matar las células diana a través de la vía perforin-granzima.
Los mastocitos (MC) también son células importantes implicadas en la inflamación en respuesta a los patógenos. Los MC actúan como células centinelas capaces de reconocer señales de peligro a través de TLR y liberan rápidamente mediadores de la inflamación tales como histamina, leucotrienos, prostaglandina, citoquinas proinflamatorias e inmunomoduladoras (IL-1, IL-3, Il-5, IL-6, GM -CSF, IL-4, IL-10, IL-13) y muchas quimiocinas. Esta actividad puede ser inducida por diferentes estímulos inflamatorios dependientes de IgE y no dependientes de IgE. Los mastocitos también pueden funcionar como células presentadoras de antígeno MHC II restringidas y expresan moléculas específicas (B7-1, B7-2, CD40) para la activación de los linfocitos T.
Los productos procedentes de parásítos y bacterias pueden ser internalizados y procesados por los mastocitos y luego presentados a los linfocitos T.
En concreto, durante las reacciones de hipersensibilidad de tipo I (alergia), los mastocitos participan en la respuesta inflamatoria, liberando mediadores cuando las IgE específicas del alergeno, unidas a receptores de inmunoglobulina E de alta afinidad (FcεRI) en su superficie celular, reaccionas a una segunda exposición al mismo alergeno.