El sistema inmunitario y la inmunidad en el cerdo: Respuesta humoral específica (2/2)
06-nov-2017 (hace 7 años 5 meses 13 días)
La respuesta humoral específica empieza cuando un linfocito B naïve reconoce su antígeno específico en los tejidos y órganos linfoides secundarios (ver "Reconocimiento de patógenos").
Cada linfocito B tiene un receptor de antígeno (BCR: Receptor de células B), una inmunoglobulina de superficie (IgM o IgD), que se une a dominios específicos del antígeno llamados determinantes antigénicos o epítopes.
La especificidad del sitio de unión antigénica depende de los genes que codifican los dominios variables (VH y VL); en los mamíferos la recombinación y/o conversión génica pueden generar un gran número de genes y, en consecuencia, muchos BCRs distintos, capacitando al animal para reconocer casi todos los antígenos externos.
Tras el reconocimiento, cada linfocito B específico se activa, realiza una proliferación clonal y se diferencia en una célula plasmática (o Célula Secretora de Anticuerpos, ASC) capaz de sintetizar y secretar inmunoglobulinas (anticuerpos), específicas para su epítope.
Las células T ayudan a optimizar la respuesta humoral específica.
Inicialmente, las células B secretan IgM en forma monomérica y con una especificidad antigénica idéntica.
En relación con el tipo de secreción de anticuerpos, los antígenos se clasifican como dependientes o independientes de células T, según si necesitan, o no, la ayuda de las células T para iniciar la respuesta de anticuerpos.
Para ciertos antígenos, los llamados independientes de células T (por ejemplo lipopolisacáridos bacterianos, flagelina, polisacáridos de pneumococos, etc.) sólo pueden inducir anticuerpos IgM con una afinidad modesta y sin células B de memoria.
Casi todos los antígenos externos son dependientes de células T y requieren, para la activación completa de los linfocitos B, una señal adicional (citoquina) procedente de linfocitos T colaboradores específicos que reconozcan al mismo antígeno (cooperación de células T y B).
En este último caso, la célula B actúa como una célula presentadora de antígenos, presentando el antígeno procesado a las células T colaboradoras que, cuando es reconocido, permite el cambio isotípico de la Ig (conmutación de clase de anticuerpo) de IgM a inmunoglobulinas de una clase distinta, IgA, IgE o IgG, dependiendo de la citoquina secretada por la célula T colaboradora, e involucradas en la estimulación de la célula B (IL-4 o IFNγ respectivamente). Otra señal básica para la conmutación del anticuerpo es la interacción del CD154 (CD40L) sobre el receptor CD40 de las células B.
Bajo esta "ayuda", las células B proliferativas experimentan una hipermutación somática de la inmunoglobulina de superficie, por lo que pasarán a estar dotadas de un isotipo específico. Estos linfocitos B son posteriormente seleccionados en base a una afinidad óptima de Ig con el antígeno al entrar en un estrecho contacto con las células dendríticas foliculares que presentan el antígeno. Sólo los linfocitos B con una elevada afinidad de anticuerpo para el antígeno, y que son capaces de procesarlo y presentarlo, serán seleccionadas positivamente. Por lo tanto sólo estos últimos linfocitos B sobrevivirán, proliferarán y se diferenciarán en células plasmáticas, sintetizando y secretando anticuerpos de una única clase isotípica, con una especificidad única y afinidad elevada, mejorando la capacidad de adherirse al antígeno y, de este modo, neutralizar y destruir los patógenos.
Cuando termina la respuesta inmune primaria, algunas de estas células B y células plasmáticas activadas y seleccionadas en este primer contacto, sobrevivirán como células de memoria de vida larga, listas para ser activadas en exposiciones posteriores al mismo antígeno (respuesta inmune secundaria) produciendo la misma clase de anticuerpos con un mayor título y afinidad.
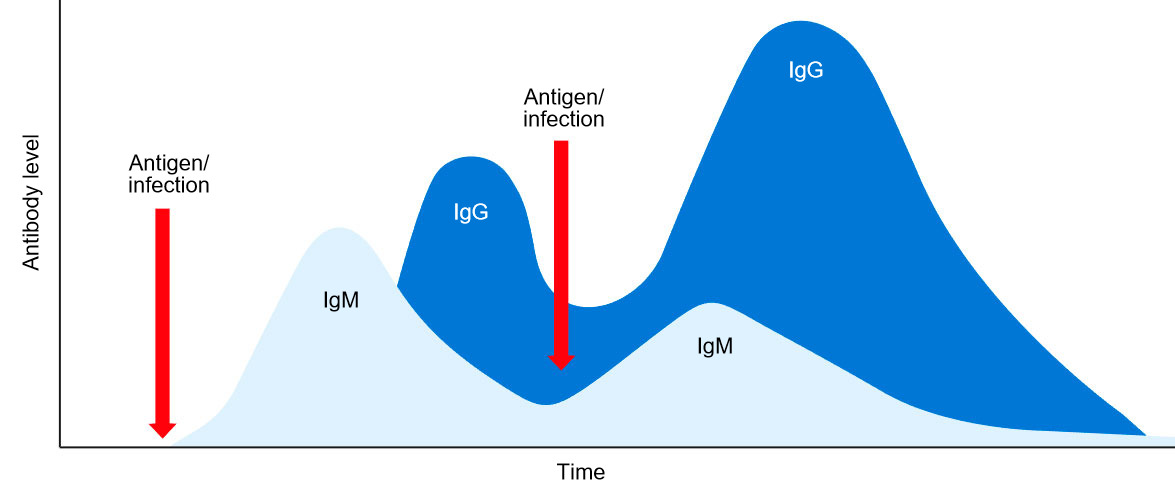
En conclusión, en la respuesta humoral primaria, se secreta inicialmente una única clase de IgM en forma monomérica y con idéntica especificidad antigénica. Luego, tras unos días, debido a la conmutación isotípica, se secretan clases distintas de Ig, por ejemplo, IgG. La respuesta humoral secundaria inducida por una exposición posterior al mismo antígeno estará caracterizada principalmente por la producción de isotipo de IgG pero con un título y afinidad más elevadas (figura 1).
Los anticuerpos pueden actuar frente a patógenos con diferentes mecanismos defensivos:
- Inhibición de la infectividad de patógenos uniéndose directamente a ellos (neutralización, bloqueo por adhesión, exclusión inmune).
- Activación de la vía alternativa (opsonización) o clásica del complemento con lisis del patógeno.
- Al recubrir al patógeno (o células infectadas), los anticuerpos permiten que las células accesorias, que reconocen porciones del fragmento cristalizable (Fc), ingieran y maten al patógeno por opsonización y fagocitosis o maten a las células infectadas mediante ADCC ("Citotoxicidad Celular dependiente de Anticuerpos").
- Regulación de la respuesta inmune a través de la interacción con los receptores Fc (FcRs).
Los FcRs se expresan en muchas células efectoras de la inmunidad innata, como los mastocitos, neutrófilos, monocitos, macrófagos y células dendríticas. Concretamente, cuando los FcγRs (receptores Fc para IgG) se unen a complejos inmunes, desencadenan la activación de las células innatas y su respuesta efectora (fagocitosis, explosión oxidativa, citotoxicidad, liberación de péptidos vasoactivos y quimioatrayentes). Los FcRs de las células dendríticas promueven la captura del antígeno, su procesado y su presentación a las células T citotóxicas y colaboradoras.
Por el contrario, la FcR inhibitoria para IgG, expresada en las células B, está implicada en la regulación de la activación y desarrollo de células B y en la supervivencia de células plasmáticas.