PRRSV en Europa: ¿de dónde viene?
09-abr-2015 (hace 9 años 11 meses 24 días)
En base a encuestas serológicas retrospectivas, la primera evidencia de PRRSV es de 1987, de la antigua República Democrática Alemana (RDA) (Ohlinger et al., 2000). En Europa se publicaron los primeros brotes clínicos en noviembre de 1990 en Alemania y, entre 1991-1992 (OIE, 1992), hubo brotes en los Países Bajos, España, Reino Unido, Francia, Bélgica y Dinamarca. Pronto, la enfermedad fue descrita en la mayoría de países europeos.
En un principio, se pensó que las cepas europeas de PRRSV (PRRSV tipo 1) están estrechamente relacionadas, pero estudios recientes han demostrado que los países de Europa del Este (Lituania, Letonia, Bielorrusia, Ucrania) y la Federación Rusa albergan cepas muy diversas de PRRSV tipo 1 (Stadejek et al., 2013). En estos países hay hasta cuatro subtipos genéticos distintos, mientras que en Europa occidental y central sólo se ha detectado un único subtipo. Este subtipo genético de Europa Occidental también está presente en América del Norte y Sureste Asiático. En Europa parece haber una marcada delimitación geográfica de la diversidad de PRRSV a lo largo de la frontera oriental de Polonia. Los virus subtipo 1 solo se detectaron al oeste de la frontera, mientras que al este están presentes los subtipos 1, 2, 3 y 4 (y probablemente más).
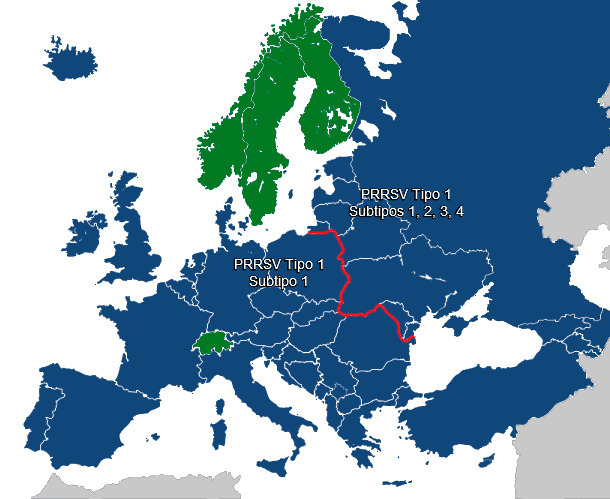
Creemos que las barreras políticas y comerciales probablemente expliquen este patrón de diversidad del virus. Las secuencias disponibles actualmente sitúan al ancestro común más reciente de las cepas de PRRSV tipo 1 entre 1946-1967, es decir, durante la posguerra de la Segunda Guerra Mundial (Forsberg, resultados no publicados), y al menos 20 años antes de los primeros signos de la presencia de este virus en Europa Occidental. En la antigua Unión Soviética, este período supuso un movimiento geográfico masivo de cerdos desde la Europa Occidental (Large White, Middle White, Danish Landrace, Berkshire), desde países incorporados a la Unión Soviética (Lithuanian y Latvian White) y desde diferentes partes de Rusia (parte asiática: Siberian, North Siberian, Siberian Black Pied, Kemerovo y Semirechensk; parte europea: Chuvash, Kuban, North Caucasus y Urzhum) dando lugar a nuevas líneas genéticas de cerdos. Parece plausible que la expansión en la posguerra de la antigua Unión Soviética y los programas de reproducción crearan un entorno que permitiera surgir un nuevo virus o propagar virus ya existente. Asimismo, dado que Polonia y otros países de Europa central estuvieron mucho menos influenciadas por estas políticas que, por ejemplo, Bielorrusia, Ucrania y los países bálticos, parece plausible que si surgieron nuevos subtipos de PRRSV durante la convulsa posguerra, estos serían dominantes en los países que antiguamente formaban parte de la Unión Soviética, pero no necesariamente en los países del antiguo bloque comunista, que no formaban parte de la Unión Soviética.
Aún así, la pregunta sobre cómo se propagó PRRSV en la parte occidental de Europa sigue abierta. PRRSV puede haberse propagado desde Alemania oriental (antigua RDA) a Europa Occidental (Ohlinger et al., 2000). Existen evidencias de que en la RDA se produjeron nuevas razas porcinas mediante el cruce con razas porcinas soviéticas. Uno de los productos resultantes de estos cruces fue el east German Large White, creado en 1954 y compuesto por German Large White, Russian Large White y Hungarian Large White con introgresión de Latvian Large White y Hungarian Yorkshire en 1980. Curiosamente esta raza de Alemania del este fue incluida oficialmente en la raza west German Large White en 1993.
La gran diversidad genética de las cepas de Europa del Este tiene implicaciones prácticas. Varios estudios independientes han demostrado que las cepas bielorrusas del subtipo genético 3 (por ejemplo, la cepa Lena) son significativamente más virulentas que cualquier otra cepa de Europa Occidental del subtipo 1 (Karniychuk et al. 2010; Morgan et al., 2013). Algunos experimentos sugieren que estas cepas pueden ser tan patógenas como las cepas de PRRS altamente patógenas procedentes de China. La diversidad genética de las cepas del Este de Europa plantea un verdadero problema para la sensibilidad de los métodos diagnósticos de PCR, ya que las regiones diana comunes de cebadores de PCR (ORF7) son genéticamente muy diversas en estas cepas, por lo que la sensibilidad de algunos de los métodos PCR de diagnóstico puede estar comprometida (Wernike et al., 2012). Además, todavía es necesario determinar la eficacia de las vacunas actuales de PRRSV contra las variantes genéticas de Europa del Este.