

La aparición de un brote de enfermedad de Glässer en una granja depende principalmente del estado inmunológico de los animales y de la virulencia de las cepas de Haemophilus parasuis circulantes.
En los primeros días de vida, los lechones reciben de sus madres tanto la bacteria como anticuerpos calostrales protectivos. Esto hace que los animales se colonicen con H. parasuis mientras están protegidos por la inmunidad materna, hasta que son capaces de producir su propia respuesta inmunitaria. El resultado es un equilibrio entre colonización y anticuerpos que mantiene las cepas bajo control y evita el desarrollo de enfermedad. La disminución del tiempo de contacto entre la madre y sus lechones, por la práctica común del destete temprano, hace que aparezcan algunos lechones que no están completamente colonizados o que no han adquirido un nivel suficiente de anticuerpos maternales. Al mezclarse con otras camadas, estos lechones se enfrentarán con cepas distintas y sin anticuerpos para poder hacer frente a la enfermedad.
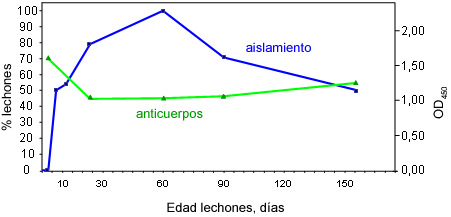

En los primeros días de vida los lechones reciben de sus madres inmunidad y bacterias. Posteriormente, ellos producen su propia respuesta. En azul, porcentaje de lechones de los cuales se aisló Haemophilus parasuis. En verde, anticuerpos (IgGs) en suero de los mismos animales.
Las cepas de H. parasuis se pueden clasificar en grupos según su serotipo (ver capítulo “Epidemiología y tipado”). Se han definido 15 serotipos en H. parasuis, pero existe además, una cantidad apreciable de cepas que no pertenecen a ninguno de estos serotipos. Cuando se definieron los 15 serotipos se realizaron infecciones experimentales con la cepa de referencia de cada serotipo y se observó una gran variabilidad de virulencia entre ellas. Desde entonces, se ha asumido que todas las cepas pertenecientes a un serotipo tienen el mismo grado de virulencia que la correspondiente cepa de referencia. En algunos casos parece probable que esta asociación sea real, como en el caso de las cepas del serotipo 5. El serotipo 5 se definió como serotipo altamente virulento y es uno de los más prevalentes, ya que por su virulencia, las cepas pertenecientes a este serotipo se aíslan frecuentemente de animales con enfermedad de Glässer. También es uno de los serotipos que más se ha utilizado para realizar infecciones experimentales, y esto ha permitido confirmar su alta virulencia.
Sin embargo hay otros datos que son más conflictivos. Nielsen, en 1993, realizó estudios de inmunidad y protección cruzada y observó que tras inoculación intranasal de cepas vivas de los serotipos 1, 2, 3, 4, 5, 6 y 7, se producían anticuerpos con reacción cruzada homóloga y heteróloga (ver Tabla). Además, los animales inoculados con los serotipos 2, 3, 4 o 7 resultaron protegidos frente a una infección intranasal con una cepa de serotipo 5. Estos resultados indican que la infección intranasal produce anticuerpos circulantes a antígenos comunes de distintos serotipos. Al contrario, en 2001 Takahashi y colaboradores, observaron que una bacterina del serotipo 2 no protegía frente a una infección intratraqueal con serotipo 5, y viceversa. Por otro lado, en 1997 Rapp-Gabrielson y colaboradores encontraron que la inmunización con bacterinas de distintos serotipos intramuscularmente, seguido de infección por vía intratraqueal, resultaba en protección variable. Cabe destacar la protección heteróloga de una bacterina combinada con serotipo 4 y 5 frente a los serotipos 13 y 14, pero no el 2 o el 12, y más sorprendentemente la falta de protección de una bacterina del serotipo 12 frente a una cepa diferente del mismo serotipo 12.
Tabla. Títulos de anticuerpos de lechones inoculados intranasalmente con cepas de referencia de distintos serotipos de Haemophilus parasuis.
| Antígeno del serotipo | |||||||
| Serotipo inoculado | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
| 1 | 64 | 0 | 16 | 0 | 0 | 0 | 0 |
| 2 | 0 | 16 | 0 | 0 | 0 | 0 | 0 |
| 3 | 32 | 0 | 16 | 0 | 0 | 0 | 0 |
| 4 | 0 | 16 | 0 | 16 | 8 | 0 | 16 |
| 5 | 0 | 16 | 0 | 8 | 16 | 16 | 8 |
| 6 | 0 | 8 | 0 | 8 | 8 | 16 | 8 |
| 7 | 0 | 16 | 0 | 8 | 8 | 8 | 16 |
| Nielsen, R. Acta Vet. Scand. 1993, 34:193-198 | |||||||
Las aparentes contradicciones de la literatura podrían ser debidas a la utilización de distintas rutas de inoculación, tanto para inmunización como para la posterior infección y a la utilización de distintas cepas dentro de un mismo serotipo, ya que también se ha observado variación en virulencia dentro de las cepas de un mismo serotipo. El serotipo está definido por las propiedades inmunológicas del antígeno soluble de las cepas (posiblemente LOS y polisacáridos), pero la composición específica de este antígeno se desconoce. El conocimiento específico de las moléculas que determinan el serotipo podría ayudar a aclarar las reacciones cruzadas entre las cepas de H. parasuis, que sería útil en establecer estrategias vacunales.
